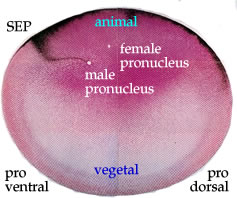
In 1980 by Kirschner, Gerhart & colleagues re-examined the process by which fertilization induces dorsal-axis in Xenopus. Their studies revealed a sperm-directed microtubule-dependent rotation of the superficial egg cortex with respect to the underlying cytoplasm. This rotation breaks the radial symmetry of the egg.
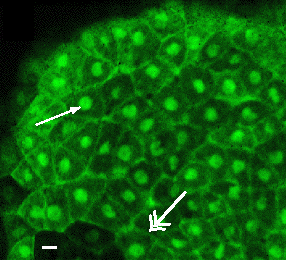
Mike Danilchick went on to characterize the complex swirls of cytoplasm that result form this rotation.
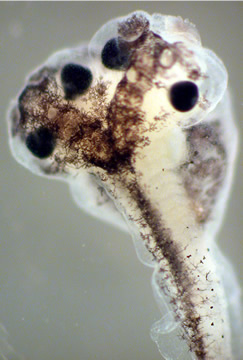
A second dorsal axis, leading to the formation of a second head, can be induced articifially by a second centrifugation-induced rearrangement.